Um umfassend über Hämoglobin (Hb) sprechen zu können, ist es nützlich, zunächst auf Myoglobin (Mb) einzugehen, das dem Hämoglobin sehr ähnlich, aber viel einfacher ist. Zwischen Hämoglobin und Myoglobin bestehen enge Verwandtschaftsbeziehungen: Beide sind konjugierte Proteine und ihre prosthetische Gruppe (Nicht-Protein-Teil) ist die Häm- Gruppe.
Myoglobin ist ein globuläres Protein, das aus einer einzelnen Kette von ungefähr 150 Aminosäuren besteht (abhängig vom Organismus) und dessen Molekulargewicht ungefähr 18 kD beträgt.
Wie erwähnt, ist es mit einer Häm-Gruppe ausgestattet, die in einen hydrophoben (oder lipophilen) Teil des Proteins eingefügt ist und aus Faltungen besteht, die auf die α-Helix-Strukturen von faserigen Proteinen zurückzuführen sind.
Myoglobin besteht hauptsächlich aus acht α-Helicesegmenten und besteht fast ausschließlich aus unpolaren Resten (Leucin, Valin, Methionin und Phenylalanin), während polare Reste praktisch nicht vorhanden sind (Asparaginsäure, Glutaminsäure, Lysin und Arginin); Die einzigen polaren Reste sind zwei Histidine, die eine grundlegende Rolle beim Angriff von Sauerstoff auf die Hämgruppe spielen.
Die Hämgruppe ist eine Chromophorgruppe (absorbiert im Sichtbaren) und ist die funktionelle Gruppe des Myoglobins.
Siehe auch: glykiertes Hämoglobin - Hämoglobin im Urin
Ein bisschen Chemie
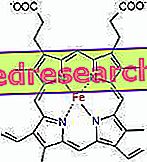
Die Verbindung zwischen Protoporphyrin und Eisen ist eine typische Bindung der sogenannten Koordinationsverbindungen, bei denen es sich um chemische Verbindungen handelt, bei denen ein zentrales Atom (oder Ion) Bindungen mit anderen chemischen Spezies eingeht, deren Anzahl höher ist als seine Oxidationszahl (elektrische Ladung). Im Falle von Häm sind diese Bindungen reversibel und schwach.
Die Koordinationszahl (Anzahl der Koordinationsbindungen) des Eisens beträgt sechs: Um das Eisen herum können sich sechs Moleküle befinden, die die Bindungselektronen teilen.
Um eine Koordinationsverbindung zu bilden, sind zwei Orbitale mit der richtigen Orientierung erforderlich: eines kann Elektronen "kaufen" und das andere kann sie abgeben
Im Häm bildet das Eisen vier planare Bindungen mit den vier Stickstoffatomen, die sich im Zentrum des Protoporphyrinrings befinden, und eine fünfte Bindung mit einem proximalen Histidinstickstoff. Eisen hat die sechste freie Koordinationsverbindung und kann an Sauerstoff binden.
Wenn Eisen in Form eines freien Ions vorliegt, haben seine d-Orbitale alle die gleiche Energie. Im Myoglobin ist das Eisenion an Protoporphyrin und Histidin gebunden. Diese Spezies stören die Orbitale des Eisens magnetisch. Das Ausmaß der Störung ist für die verschiedenen Orbitale je nach ihrer räumlichen Ausrichtung und der der störenden Arten unterschiedlich. Da die Gesamtenergie der Orbitale wieder konstant sein muss, bewirkt die Störung eine energetische Trennung zwischen den verschiedenen Orbitalen: Die von einigen Orbitalen aufgenommene Energie entspricht dem Energieverlust der anderen.
Wenn der Abstand zwischen den Orbitalen nicht sehr groß ist, ist eine elektronische Anordnung mit hohem Spin vorzuziehen: Die Bindungselektronen versuchen, sich in mehreren möglichen Unterebenen in parallelen Spins anzuordnen (maximale Multiplizität); Wenn andererseits die Störung sehr stark ist und ein großer Abstand zwischen den Orbitalen besteht, ist es möglicherweise zweckmäßiger, die Bindungselektronen in den Orbitalen mit niedrigerer Energie zu paaren (niedriger Spin).
Wenn Eisen an Sauerstoff bindet, nimmt das Molekül eine geringe Spinneigung an, während das Molekül bei Eisen mit der sechsten freien Koordinationsverbindung einen hohen Spin aufweist.
Dank dieses Unterschieds im Spin können wir durch eine Spektralanalyse von Myoglobin verstehen, ob Sauerstoff gebunden ist (MbO2) oder nicht (Mb).
Myoglobin ist ein muskeltypisches Protein (das aber nicht nur in den Muskeln vorkommt).
Myoglobin wird aus dem Pottwal gewonnen, in dem es in großen Mengen vorhanden ist, und anschließend gereinigt.
Wale atmen wie Menschen: Sie haben die Lunge und müssen über die Atemwege Luft aufnehmen. Der Pottwal muss so viel Sauerstoff wie möglich in den Muskeln haben, die in der Lage sind, Sauerstoff anzusammeln, indem sie ihn an das in ihnen vorhandene Myoglobin binden. Sauerstoff wird dann langsam freigesetzt, wenn der Wal eingetaucht wird, weil sein Stoffwechsel Sauerstoff erfordert: Je mehr Sauerstoff der Pottwal aufnehmen kann und je mehr Sauerstoff während des Tauchgangs verfügbar ist.

Das Fleisch mehr oder weniger rot zu machen, ist der Gehalt an Hämoproteinen (es ist das Häm, das das Fleisch rot macht).
Hämoglobin hat viele strukturelle Analogien zu Myoglobin und ist in der Lage, molekularen Sauerstoff reversibel zu binden. Während Myoglobin im Allgemeinen auf Muskeln und peripheres Gewebe beschränkt ist, findet sich Hämoglobin in Erythrozyten oder roten Blutkörperchen (es handelt sich um Pseudozellen, das heißt, es handelt sich nicht um echte Zellen), die 40% des Blutes ausmachen.
Hämoglobin ist ein Tetrameter, dh es besteht aus vier Polypeptidketten mit jeweils einer Hämgruppe und zwei mal zwei (in einem Menschen gibt es zwei Alpha- und zwei Betaketten).
Die Hauptfunktion des Hämoglobins ist der Sauerstofftransport ; Eine weitere Funktion des Blutes, an dem Hämoglobin beteiligt ist, ist der Transport von Substanzen zu den Geweben.
Auf dem Weg von der sauerstoffreichen Lunge zum Gewebe transportiert das Hämoglobin Sauerstoff (gleichzeitig gelangen auch andere Stoffe in das Gewebe), während es auf dem umgekehrten Weg den aus dem Gewebe gesammelten Abfall mit sich führt, insbesondere das im Stoffwechsel produzierte Kohlendioxid.
In der Entwicklung eines Menschen gibt es Gene, die nur für einen bestimmten Zeitraum exprimiert werden; Aus diesem Grund haben wir verschiedene Hämoglobine: fötale, embryonale des erwachsenen Mannes.
Die Ketten, aus denen diese verschiedenen Hämoglobine bestehen, haben unterschiedliche Strukturen, aber bei Ähnlichkeiten ist die Funktion, die sie erfüllen, mehr oder weniger gleich.
Eine Erklärung für das Vorhandensein mehrerer verschiedener Ketten lautet wie folgt: Während des Evolutionsprozesses von Organismen hat sich auch Hämoglobin entwickelt, das sich auf den Sauerstofftransport aus Gebieten spezialisiert hat, die reich an Mangelgebieten sind. Zu Beginn der Evolutionskette beförderte das Hämoglobin Sauerstoff in kleinen Organismen; Im Laufe der Evolution haben die Organismen größere Dimensionen erreicht, daher hat sich das Hämoglobin verändert, um Sauerstoff in Gebiete zu transportieren, die weiter von dem Punkt entfernt sind, an dem er reich ist. Dazu wurden im Verlauf des Evolutionsprozesses neue Strukturen der Ketten, die das Hämoglobin bilden, kodifiziert.
Myoglobin bindet Sauerstoff auch bei geringem Druck; In peripheren Geweben herrscht ein Druck (PO2) von etwa 30 mmHg: Bei diesem Druck setzt Myoglobin keinen Sauerstoff frei, so dass es als Sauerstoffträger unwirksam wäre. Hämoglobin hingegen hat ein elastischeres Verhalten: Es bindet Sauerstoff bei hohem Druck und setzt ihn frei, wenn der Druck abnimmt.
Wenn ein Protein funktionell aktiv ist, kann es seine Form leicht verändern. Beispielsweise hat sauerstoffhaltiges Myoglobin eine andere Form als nicht sauerstoffhaltiges Myoglobin, und diese Mutation betrifft benachbarte nicht.
Bei assoziierten Proteinen wie Hämoglobin ist der Diskurs anders: Wenn eine Kette mit Sauerstoff angereichert wird, wird sie dazu veranlasst, ihre Form zu ändern, aber diese Modifikation ist dreidimensional, daher sind auch die anderen Ketten des Tetrameters betroffen. Die Tatsache, dass die Ketten miteinander assoziiert sind, lässt vermuten, dass die Modifikation einer Kette andere benachbarte beeinflusst, wenn auch in unterschiedlichem Ausmaß; Wenn eine Kette mit Sauerstoff angereichert ist, nehmen die anderen Ketten des Tetrameters eine "weniger feindliche Haltung" gegenüber Sauerstoff ein: Die Schwierigkeit, mit der eine Kette mit Sauerstoff angereichert wird, nimmt ab, wenn sich die Ketten neben ihr selbst mit Sauerstoff angereichert haben. Das gleiche Argument gilt für die Sauerstoffentfernung.
Die quaternäre Struktur von Desoxyhämoglobin wird als T- Form (Tense-Form) bezeichnet, während die von Oxyhämoglobin als R- Form (Release-Form) bezeichnet wird. Im gestreckten Zustand gibt es eine Reihe ziemlich starker elektrostatischer Wechselwirkungen zwischen sauren Aminosäuren und basischen Aminosäuren, die zu einer starren Struktur von Desoxyhämoglobin (deshalb die "angespannte Form") führen, während, wenn Sauerstoff gebunden ist, die Einheit von diesen ist Wechselwirkungen nehmen ab (deshalb die "freigegebene Form"). Darüber hinaus wird in Abwesenheit von Sauerstoff die Histidinladung (siehe Struktur) durch die entgegengesetzte Ladung von Asparaginsäure stabilisiert, während in Gegenwart von Sauerstoff das Protein dazu neigt, ein Proton zu verlieren; All dies bedeutet, dass sauerstoffhaltiges Hämoglobin eine stärkere Säure als sauerstoffarme Hämoglobie ist: Bohr-Effekt .
Abhängig vom pH-Wert bindet die Hämgruppe mehr oder weniger leicht an Sauerstoff: In einer sauren Umgebung setzt das Hämoglobin Sauerstoff leichter frei (die angespannte Form ist stabil), während in einer basischen Umgebung die Bindung mit Sauerstoff besteht stärker.

Jedes Hämoglobin setzt 0, 7 Protonen pro Mol Sauerstoff (O2) frei.
Durch den Bohr-Effekt kann Hämoglobin seine Fähigkeit verbessern, Sauerstoff zu transportieren.
Das Hämoglobin, das den Weg von der Lunge zum Gewebe zurücklegt, muss sich nach Druck, pH-Wert und Temperatur ausgleichen.
Wir sehen den Einfluss der Temperatur .
Die Temperatur in den Lungenalveolen ist etwa 1-1, 5 ° C niedriger als die Außentemperatur, während in den Muskeln die Temperatur etwa 36, 5-37 ° C beträgt; Mit steigender Temperatur sinkt der Sättigungsfaktor (bei gleichem Druck): Dies geschieht, weil die kinetische Energie zunimmt und die Dissoziation begünstigt wird.

Es gibt andere Faktoren, die die Fähigkeit von Hämoglobin, sich an Sauerstoff zu binden, beeinflussen können. Einer davon ist die Konzentration von 2, 3-Bisphosphoglycerat .
2, 3-Bisphosphoglycerat ist ein Stoffwechselmittel, das in Erythrozyten in einer Konzentration von 4 bis 5 mM vorhanden ist (in keinem anderen Körperteil ist es in einer derart hohen Konzentration vorhanden).
Bei physiologischem pH-Wert ist 2, 3-Bisphosphoglycerat deprotoniert und weist fünf negative Ladungen auf. es wird zwischen den beiden Beta-Ketten des Hämoglobins eingeklemmt, da diese Ketten eine hohe Konzentration positiver Ladungen aufweisen. Die elektrostatischen Wechselwirkungen zwischen den Beta-Ketten und den Bisphosphoglyceratketten verleihen dem System eine gewisse Steifheit: Man erhält eine gespannte Struktur mit geringer Affinität zu Sauerstoff; während der Oxygenierung wird dann 2, 3-Bisphosphoglycerat ausgestoßen.
In den Erythrozyten gibt es einen speziellen Apparat, der 1, 3-Bisphosphoglycerat (produziert vom Metabolismus) in 2, 3-Bisphosphoglycerat umwandelt, so dass es eine Konzentration von 4 bis 5 mM erreicht und das Hämoglobin daher in der Lage ist, l auszutauschen Sauerstoff im Gewebe.
Das an einem Gewebe ankommende Hämoglobin befindet sich im freigesetzten Zustand (an Sauerstoff gebunden), aber in der Nähe des Gewebes ist es carboxyliert und geht in einen gedehnten Zustand über: Das Protein hat in diesem Zustand eine geringere Tendenz, sich an Sauerstoff zu binden. im Vergleich zum freigesetzten Zustand gibt das Hämoglobin daher Sauerstoff an das Gewebe ab; außerdem entstehen durch die reaktion zwischen wasser und kohlendioxid h + ionen und somit durch den bohr-effekt sauerstoff.
Kohlendioxid diffundiert durch die Plasmamembran in den Erythrozyten. Angesichts der Tatsache, dass Erythrozyten etwa 40% des Blutes ausmachen, ist zu erwarten, dass nur 40% des Kohlendioxids, das aus den Geweben diffundiert, in die Erythrozyten gelangen, da sie ein Enzym enthalten, das sich umwandelt Aus Kohlendioxid in Kohlensäure folgt, dass die stationäre Kohlendioxidkonzentration in den Erythrozyten niedrig ist und daher die Eintrittsgeschwindigkeit hoch ist.

Ein anderes Phänomen, das auftritt, wenn ein Erythrozyten ein Gewebe erreicht, ist das folgende: Durch einen Gradienten tritt das HCO3- (Kohlendioxid-Derivat) aus dem Erythrozyten aus, und um den Austritt einer negativen Ladung auszugleichen, haben wir das Eintritt von Chloriden, die einen Anstieg des osmotischen Drucks bestimmen: Um diese Schwankung auszugleichen, tritt auch Wasser ein, das eine Erythrozytenschwellung verursacht (HAMBURGER-Effekt). Das gegenteilige Phänomen tritt auf, wenn ein Erythrozyten die Lungenbläschen erreicht: Es tritt eine Erythrozyten-Deflation auf (HALDANE-Effekt). Somit sind venöse Erythrozyten (direkt zur Lunge) runder als arterielle.